Кратко
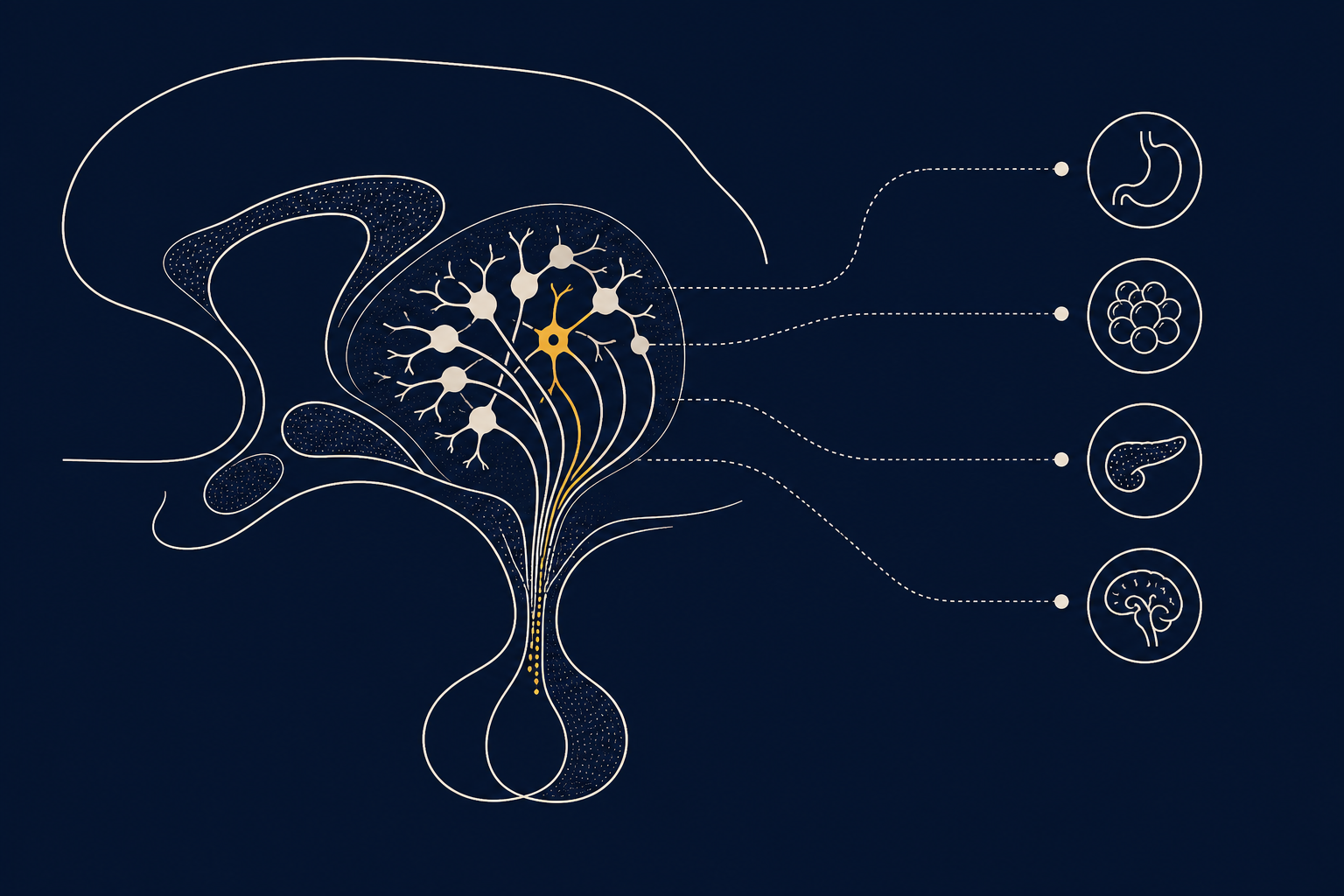
Аркуатное ядро гипоталамуса (Arcuate Nucleus, ARC) — небольшое скопление нейронов у основания третьего желудочка мозга, одна из главных сенсорных и командных станций энергетического и гормонального баланса. Отсюда берёт начало тубероинфундибулярный дофаминовый путь: специализированная группа дофаминовых нейронов A12 тонически подавляет секрецию пролактина гипофизом. В том же ядре расположены нейроны, запускающие голод и насыщение, поэтому ARC часто называют «воротами» между сигналами тела и мотивационными цепями мозга.
Что это
Аркуатное ядро лежит в медиобазальном гипоталамусе, у дна третьего желудочка, вплотную к срединному возвышению — зоне с ослабленным гематоэнцефалическим барьером. Такое соседство неслучайно: сюда легко проникают циркулирующие в крови гормоны и метаболиты — лептин, инсулин, глюкоза, грелин, — и именно здесь центральная нервная система впервые «считывает» их. В ARC совмещены функционально разные, но топографически перекрывающиеся популяции клеток: орексигенные AgRP/NPY-нейроны и анорексигенные POMC-нейроны, задающие баланс голода и сытости, и дофаминовые нейроны A12, определяющие уровень пролактина. Тубероинфундибулярная дофаминовая (TIDA) система — единственный дофаминовый путь мозга, выполняющий не психическую, а строго эндокринную функцию: он связывает гипоталамус с передней долей гипофиза.
Как это работает
Тела дофаминовых нейронов TIDA лежат в аркуатном ядре, а их аксоны заканчиваются в наружной зоне срединного возвышения, откуда выделившийся дофамин попадает в портальные сосуды гипофиза и связывается с D2-рецепторами лактотрофов, подавляя выброс пролактина. Это классическая петля отрицательной обратной связи по короткому контуру: рост уровня пролактина в крови сам стимулирует TIDA-нейроны увеличивать синтез и выброс дофамина, а тот гасит дальнейшую секрецию гормона. Активность этих клеток колеблется фазно-тонически и зависит от эстрогенов, опиоидов, окситоцина, а также моноаминергических волокон — например, норадренергических, действующих в том числе через α2A-адренорецепторы и тонко подстраивающих возбудимость нейронов под текущее состояние организма. Доступ подобных сигналов к клеткам ARC во многом контролируют астроциты и родственные им танициты — глия на границе срединного возвышения, физически и метаболически определяющая, какие молекулы крови допускаются к телам нейронов.
Почему это важно для мотивации
На первый взгляд регуляция пролактина далека от темы усилия и вознаграждения. Но аркуатное ядро — узел, где метаболический сигнал буквально конвертируется в мотивационное состояние. Голод, зафиксированный AgRP-нейронами ARC, не просто вызывает желание поесть: он системно смещает то, как мозг взвешивает выгоды и издержки любого действия. Активация этих нейронов резко меняет поведенческие приоритеты — организм готов прилагать больше усилий и принимать больше риска ради еды, то есть меняется сама логика анализа затрат и выгод. Схожим образом смещается и терпимость к отсрочке: в состоянии голода мозг систематически предпочитает менее ценную, но немедленную награду отложенной и большей — эффект, тесно связанный с аверсией к задержке. Дофаминовая ветвь ARC добавляет к этой картине эндокринный контур: пролактин, секреция которого определяется активностью TIDA-нейронов, модулирует половое поведение и чувствительность дофаминовых цепей вознаграждения, а хроническая гиперпролактинемия на фоне некоторых препаратов часто сопровождается снижением либидо и общего мотивационного тонуса.
Что с этим делать
Для практики важны два вывода. Первый: устойчивый, предсказуемый режим питания снижает амплитуду голодных сигналов из ARC и смягчает связанные с голодом искажения решений — импульсивность, тягу к немедленному вознаграждению, недооценку долгосрочных выгод. Принимать решения «на пустой желудок» невыгодно именно потому, что аркуатное ядро в этот момент перестраивает расчёт цены усилия. Второй: если на фоне приёма дофамин-блокирующих препаратов или других вмешательств появляются признаки гиперпролактинемии — снижение либидо, нарушения цикла, нетипичная лактация, — стоит помнить, что причина может крыться в подавлении TIDA-дофамина, а не в психологии, и обсудить это с врачом.
Итог
Аркуатное ядро гипоталамуса — компактная, но функционально двойная структура: с одной стороны, источник дофаминовых нейронов A12, тонически подавляющих пролактин по механизму короткой обратной связи; с другой — центр детекции голода и энергетического баланса, задающий фон для мотивационных вычислений мозга. Обе роли объединяет одна логика: ARC переводит физиологические сигналы тела в нейрохимические команды, определяющие и гормональный статус, и то, ради чего мозг готов действовать.
Материал носит образовательный характер и не заменяет консультацию специалиста.