Кратко
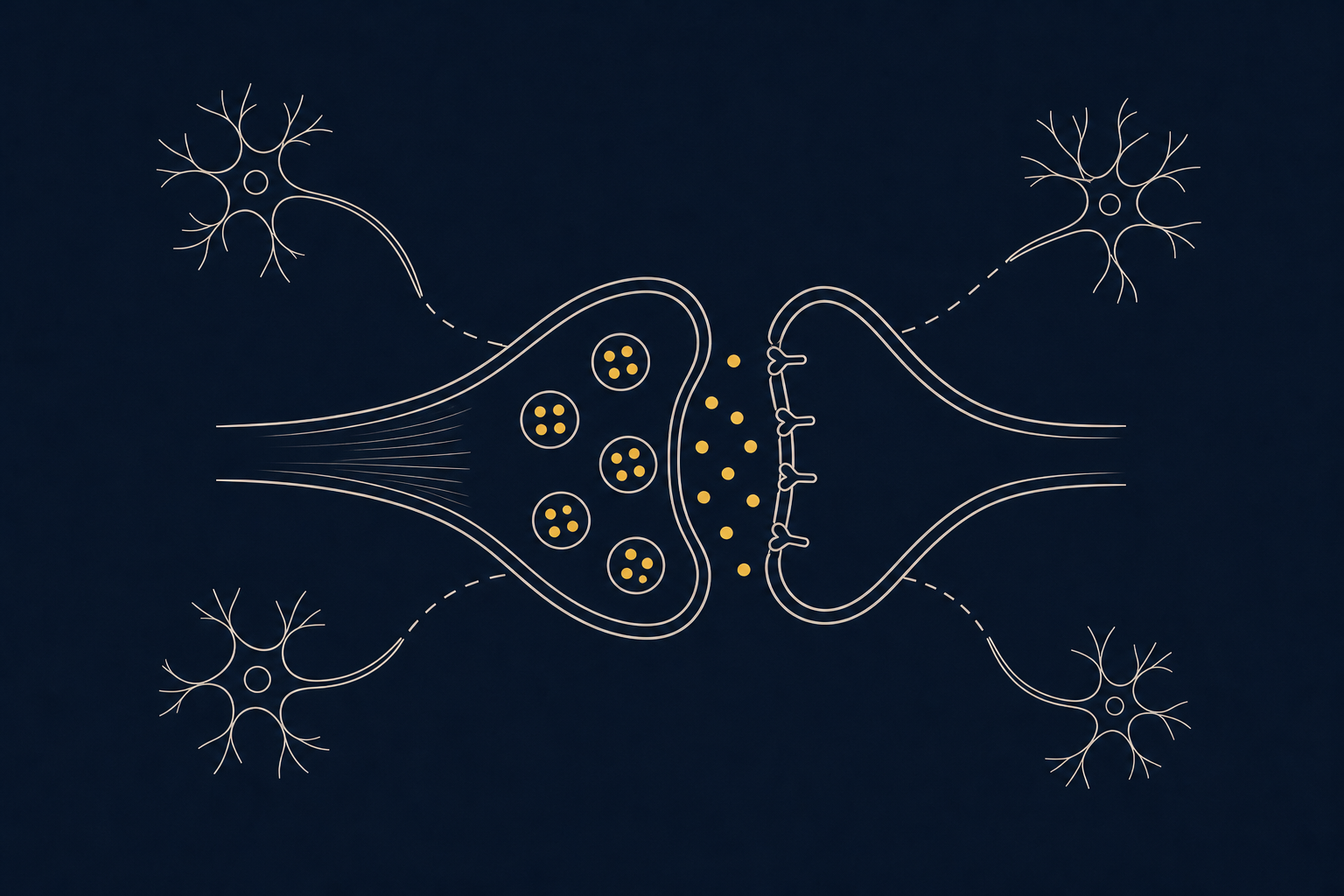
5-HT2C-рецептор — один из подтипов серотониновых рецепторов, который в вентральной области покрышки (ВОП) среднего мозга работает как тормозной клапан для дофаминовой системы. При активации он возбуждает соседние ГАМК-нейроны, а те подавляют дофаминовые клетки, снижая выброс дофамина в прилежащем ядре. Именно поэтому этот рецептор так интересен исследователям мотивации, импульсивности и зависимостей: усиление его работы ослабляет стремление к вознаграждению, а блокада, наоборот, его усиливает.
Что это
Серотониновые рецепторы делятся на несколько семейств — 5-HT1, 5-HT2, 5-HT3 и другие, — и 5-HT2C относится ко второму, метаботропному типу, сопряжённому с G-белком Gq/11. В отличие от близкого по структуре 5-HT2A-рецептора, который сосредоточен в коре и связан с действием психоделиков, 5-HT2C плотно представлен в лимбических структурах — сосудистом сплетении, миндалине, гиппокампе, а главное для темы мотивации — в вентральной области покрышки и прилежащем ядре, ключевых узлах мезолимбического дофаминового пути. Рецептор кодируется геном HTR2C на X-хромосоме, а его РНК подвергается редактированию, поэтому в мозге одновременно существует несколько функциональных изоформ белка с разной эффективностью передачи сигнала.
Как это работает
Иммуногистохимические исследования показывают, что в ВОП 5-HT2C-рецепторы находятся в основном на ГАМК-ергических нейронах и в меньшей степени — непосредственно на дофаминовых клетках. Когда серотонин связывается с рецептором на ГАМК-нейроне, тот возбуждается и усиливает торможение соседних дофаминовых клеток. В итоге падает выброс дофамина в прилежащем ядре — этот трёхзвенный контур «серотонин → 5-HT2C-рецептор → ГАМК-нейрон → дофаминовый нейрон ВОП» и лежит в основе тормозного влияния серотонина на мотивационную дофаминовую систему. Небольшая часть 5-HT2C-рецепторов расположена и на самих дофаминовых нейронах, что позволяет тонко балансировать силу эффекта.
Похожая логика действует и в прилежащем ядре, где 5-HT2C-рецепторы находятся на ГАМК-эфферентных нейронах и модулируют локальные тормозные петли, а также обратную связь на ВОП. Поэтому конечный эффект стимуляции рецептора зависит от того, в какой именно структуре он преимущественно активируется — иногда эффекты в разных отделах мезолимбического пути оказываются даже противоположными.
Почему это важно для мотивации
Дофаминовые нейроны ВОП кодируют не «удовольствие» сам по себе, а ожидаемую ценность усилия и вероятность вознаграждения — то, что толкает поведение вперёд. Поскольку 5-HT2C-рецептор способен приглушать этот сигнал, он становится естественным тормозом побуждения. В экспериментах с прогрессивным коэффициентом подкрепления, где для получения награды нужно прилагать всё больше усилий, стимуляция 5-HT2C-рецепторов ВОП снижала готовность животных работать за вознаграждение — то есть напрямую уменьшала мотивацию к усилию, а не просто вызывала сытость или седацию. Похожие данные получены и для эффектов психостимуляторов: активация 5-HT2C в ВОП ослабляет дофаминовый ответ на кокаин. Это помогает понять, почему сниженный серотониновый тонус (например, у носителей короткого варианта гена переносчика серотонина, см. 5-HTTLPR) связывают с усилением импульсивности и рискового поведения: слабее тормозной контур — сильнее нерегулируемый дофаминовый порыв. Взаимодействие медиаторных систем здесь устроено как система сдержек и противовесов: похожую тормозную роль по отношению к дофаминовому усилию играет и A2A-рецептор аденозина в стриатуме, хотя механизм у него принципиально иной.
Что с этим делать
5-HT2C-рецептор — не переключатель силы воли, а часть биохимического баланса, который стоит учитывать, а не пытаться «взломать». Практический вывод касается в первую очередь фармакологии и самонаблюдения, а не самостоятельной коррекции поведения. Агонисты 5-HT2C в разное время исследовались как средства для снижения тяги к еде и никотину именно за счёт торможения дофаминового вознаграждения — это иллюстрирует, что избыточная стимуляция рецептора снижает не только импульсивные порывы, но и здоровую целенаправленную мотивацию. Обратная сторона: препараты и состояния, снижающие серотониновый тонус, потенциально расторможивают дофаминовый контур и повышают импульсивность. Если готовность прилагать усилие резко падает на фоне серотонинергических препаратов (антидепрессанты, средства для контроля веса) или растёт импульсивность на фоне их отмены, разумнее обсудить это с врачом, а не списывать на характер.
Итог
5-HT2C-рецептор — один из главных «тормозов» мезолимбической дофаминовой системы: через контур с участием ГАМК-нейронов ВОП он снижает выброс дофамина и тем самым ослабляет мотивацию к усилию и импульсивные порывы. Его работа объясняет часть индивидуальных различий в готовности работать за вознаграждение и в устойчивости к соблазнам, а вместе с другими участниками мотивационной химии, такими как DRD2 Taq1A/ANKK1, формирует общий баланс между «хочу» и «сдерживаюсь».
Материал носит образовательный характер и не заменяет консультацию специалиста.