Кратко
Аденозинтрифосфат (АТФ) — универсальная молекула-посредник, которая накапливает и передаёт энергию во всех клетках, включая нейроны. Любое действие мозга — от потенциала действия до синтеза белка — оплачивается гидролизом АТФ. Когда молекула отрабатывает свой ресурс, она распадается до аденозина — вещества, которое, в отличие от самого АТФ, обладает собственной сигнальной функцией: тормозит возбуждение, регулирует сон и модулирует дофаминовую систему, то есть напрямую связано с ощущением усилия и готовностью действовать.
Что это
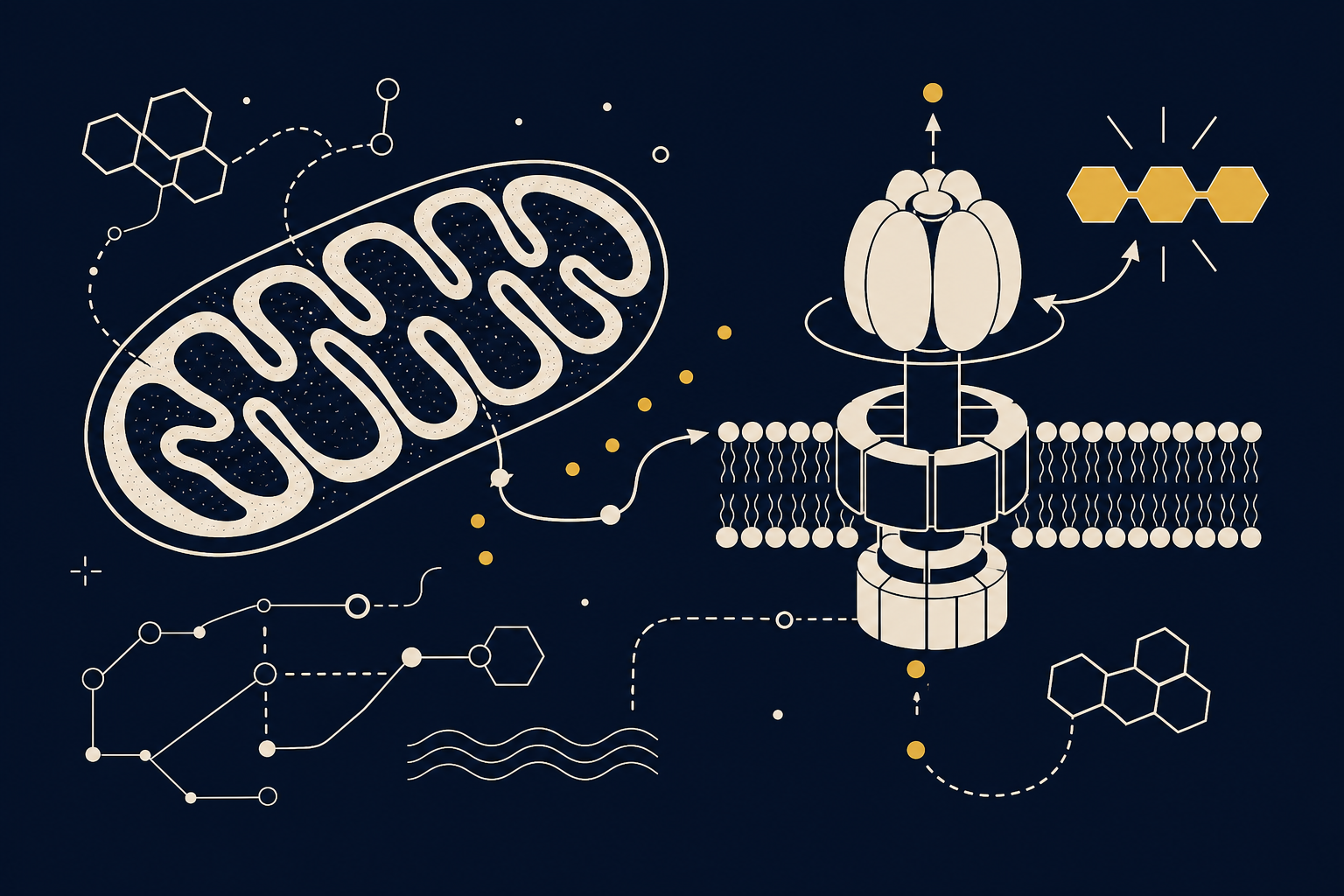
АТФ (Adenosine Triphosphate) состоит из аденина, сахара рибозы и трёх остатков фосфорной кислоты. Связи между вторым и третьим фосфатами высокоэнергетические: их разрыв высвобождает энергию для механической работы, транспорта веществ через мембраны, синтеза макромолекул и передачи сигналов. Поэтому биохимики называют АТФ «энергетической валютой клетки» — она не откладывается про запас, а непрерывно тратится и восполняется, как оборотные средства предприятия. Для нервной ткани это особенно критично: мозг составляет около 2% массы тела, но потребляет порядка 20% энергии всего организма, и большая часть этих затрат уходит на поддержание ионных градиентов через натрий-калиевый насос.
Как это работает
Основной объём АТФ синтезируется в митохондриях в процессе окислительного фосфорилирования. Питер Митчелл в 1961 году показал, что перенос электронов по дыхательной цепи создаёт градиент протонов на внутренней мембране митохондрии, а обратный поток протонов через фермент АТФ-синтазу вращает её ротор и «сшивает» АДФ с фосфатом — эта хемиосмотическая теория до сих пор остаётся основой представлений о клеточной энергетике. Использованная молекула АТФ теряет фосфаты последовательно: сначала образуется АДФ, затем АМФ, а на последнем шаге фермент 5'-нуклеотидаза отщепляет последний фосфат, оставляя аденозин — нуклеозид, который выходит за пределы клетки и действует уже не как источник энергии, а как самостоятельный сигнал.
Кроме внутриклеточного метаболизма, АТФ выделяется во внеклеточное пространство и работает как медиатор пуринергической системы: астроциты выбрасывают АТФ путём кальций-зависимого экзоцитоза, а внеклеточные ферменты быстро гидролизуют его до аденозина, который действует на рецепторы A1 и A2A соседних нейронов. Этим механизмом объясняется накопление «давления сна»: чем дольше бодрствование, тем выше внеклеточный аденозин и тем сильнее его тормозящее влияние на кору.
Почему это важно для мотивации
Связь между энергетическим обменом и мотивацией не метафорична, а физиологична. Аденозин, продукт распада АТФ, через рецепторы A2A образует функциональные комплексы с дофаминовыми D2-рецепторами в прилежащем ядре: активация A2A ослабляет сигнал D2, и наоборот. В экспериментах агонисты A2A воспроизводят эффект дофаминового дефицита — животные выбирают менее трудозатратные варианты вознаграждения, тогда как антагонисты A2A возвращают готовность прилагать усилие даже при блокаде D2-рецепторов. Это одна из причин, почему накопление аденозина на фоне усталости и недосыпа ощущается как снижение мотивации и растущая аверсия к задержке результата: система предпочитает быстрый и лёгкий выигрыш, экономя энергию. Действие кофеина — самого массового психостимулятора — построено на блокаде аденозиновых рецепторов, что временно снимает их тормозящее влияние на дофамин и создаёт ощущение прилива сил. У людей с депрессией меняется митохондриальная биоэнергетика: клетки работают на пределе в покое, но хуже справляются с ростом нагрузки, и эта сниженная резервная мощность коррелирует с усталостью и мотивационным дефицитом.
Что с этим делать
Практический вывод не в том, чтобы искусственно «повышать уровень АТФ» — это невозможно и не нужно, а в том, чтобы учитывать энергетическую логику мозга при объяснении спадов мотивации. Хроническое недосыпание и монотонная нагрузка увеличивают внеклеточный аденозин и, как следствие, ощущение бессилия и нежелания действовать — и это обратимо через полноценный сон, а не волевое усилие. Скорость обратного клиренса аденозина регулируют ферменты аденозиндезаминаза и аденозинкиназа, поэтому индивидуальные различия в переносимости усталости и чувствительности к кофеину частично объясняются активностью именно этих ферментов, а не «слабой волей». Устойчивость к усилию ограничена реальной клеточной энергетикой, и восстановление — не роскошь, а часть работы с мотивацией.
Итог
АТФ — не просто формула из школьного курса биологии, а материальная основа любого произвольного действия и, через свой метаболит аденозин, один из регуляторов готовности действовать. Цепочка от митохондрии до дофаминового рецептора объясняет, почему усталость снижает мотивацию не только «психологически», но и на уровне измеримой биохимии, и почему борьба с прокрастинацией иногда начинается не с дисциплины, а с восстановления энергетического баланса организма.
Материал носит образовательный характер и не заменяет консультацию специалиста.