Кратко
Ацетилхолин (АХ, англ. Acetylcholine, ACh) — один из первых открытых нейромедиаторов и один из самых многозначных: в коре и таламусе он настраивает внимание и обучение, в базальных ганглиях — модулирует ответ дофаминовых нейронов на награду, а на периферии управляет мышцами и вегетативной нервной системой. Для мотивации важна именно центральная, «настроечная» роль АХ: он определяет, насколько мозг готов замечать новое и придавать вес тому, что было получено усилием.
Что это
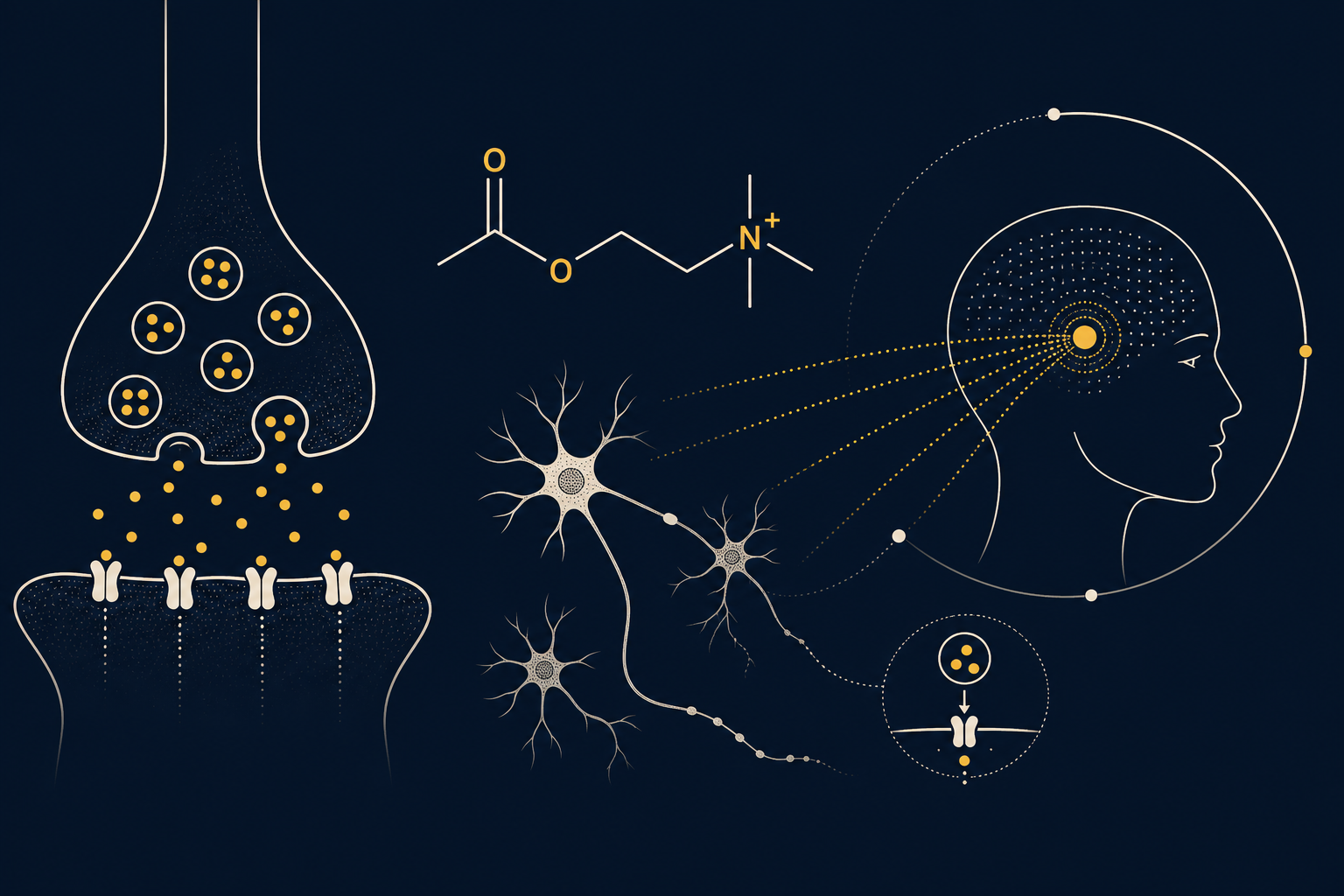
Ацетилхолин синтезируется из холина и ацетил-КоА ферментом холинацетилтрансферазой и разрушается ацетилхолинэстеразой — поэтому его действие в синапсе короткое и точное. В нейрохимии мотивации АХ занимает промежуточное положение между «быстрыми» медиаторами вроде глутамата и «медленными» модуляторами вроде дофамина: он не запускает движение напрямую, а меняет режим работы целых сетей, переключая кору между состояниями «сбора информации» и «использования уже накопленного опыта» (Hasselmo & McGaughy, 2004). Основные источники центрального АХ — базальное ядро Мейнерта, посылающее проекции в кору и миндалину, и стриатальные холинергические интернейроны, локально регулирующие дофаминовую передачу в полосатом теле.
Как это работает
Значимы два холинергических контура. Первый — кортикальный: аксоны из базального ядра Мейнерта выделяют АХ диффузно по коре, усиливая ответ на приходящий сенсорный сигнал и подавляя внутреннюю, «воспоминательную» активность — именно этот механизм повышает внимание и точность кодирования новой информации (Sarter & Bruno, 1997; Hasselmo, 2006). Второй — стриатальный: крупные холинергические интернейроны, тонически активные нейроны полосатого тела, синхронно замолкают на 100–200 мс в ответ на значимые или неожиданные стимулы. Эта пауза снимает тормозящее влияние АХ на никотиновые рецепторы дофаминовых окончаний и позволяет фазическому выбросу дофамина точнее «достучаться» до постсинаптических клеток — то есть усиливает контраст сигнала об ошибке предсказания награды (Cragg, 2006). Подробнее о том, как эта холинергическая пауза тормозит и переключает контуры внимания, — в статье про «аксональный тормоз» холинергии.
Почему это важно для мотивации
АХ и дофамин работают в связке, а не по отдельности. Пауза холинергических интернейронов синхронизирована со всплеском дофаминовой активности, и именно момент «затишья» АХ во многом определяет, будет ли награда воспринята мозгом как значимая и заслуживающая закрепления в памяти о ценности действия (Picciotto, Higley & Mineur, 2012). Если холинергическая настройка нарушена — избыточна или недостаточна, — дофаминовый сигнал о награде «смазывается»: субъективная ценность усилия и заработанного результата считывается менее отчётливо, внимание рассеивается, а обучение на успехах и неудачах замедляется. Кортикальный АХ дополнительно взаимодействует с норадренергической системой (см. α2A-адренорецепторы) и с аденозиновой регуляцией бодрствования (см. аденозин): вместе эти системы формируют общий тонус готовности прикладывать усилие и терпеть отсрочку награды, о чём подробнее в материале про аверсию к задержке.
Что с этим делать
Практические выводы из холинергической модели мотивации скромны, но конкретны. Устойчивое внимание и качественное усвоение новой информации требуют условий, где АХ может выполнять свою «настроечную» работу: многозадачность и хронический недосып подавляют холинергический тонус базального ядра Мейнерта, из-за чего обучение страдает быстрее, чем кажется на первый взгляд. Поскольку холинергическая пауза помогает мозгу «заметить» награду, задача, разбитая на понятные, различимые этапы с явными точками завершения, даёт более чёткие сигналы для дофаминовой системы, чем расплывчатый непрерывный процесс. Наконец, возрастное и патологическое снижение числа холинергических нейронов базального ядра — один из ранних маркеров нейродегенерации, поэтому стойкие жалобы на внимание и мотивацию у пожилых людей требуют медицинской, а не только поведенческой оценки (Mesulam et al., 2013).
Итог
Ацетилхолин — не «медиатор мотивации» в прямом смысле, а система точной настройки, которая решает, когда мозгу выгодно внимательно смотреть на мир, а когда — использовать то, что он уже знает, и которая помогает дофаминовым сигналам о награде оставаться различимыми и точными. Именно поэтому нарушения холинергической регуляции сказываются не столько на желании действовать напрямую, сколько на способности замечать, ценить и закреплять результат усилия.
Материал носит образовательный характер и не заменяет консультацию специалиста.