Кратко
D1-рецептор — один из пяти типов дофаминовых рецепторов, самый распространённый в мозге представитель D1-подобного семейства. Он не тормозит, а возбуждает нейрон: связываясь с дофамином, рецептор запускает G-белок Gs, повышает уровень цАМФ и меняет возбудимость клетки. Этот механизм лежит в основе рабочей памяти, целенаправленного поведения и — при избыточной или недостаточной активации — сбоев когнитивного контроля.
Что это
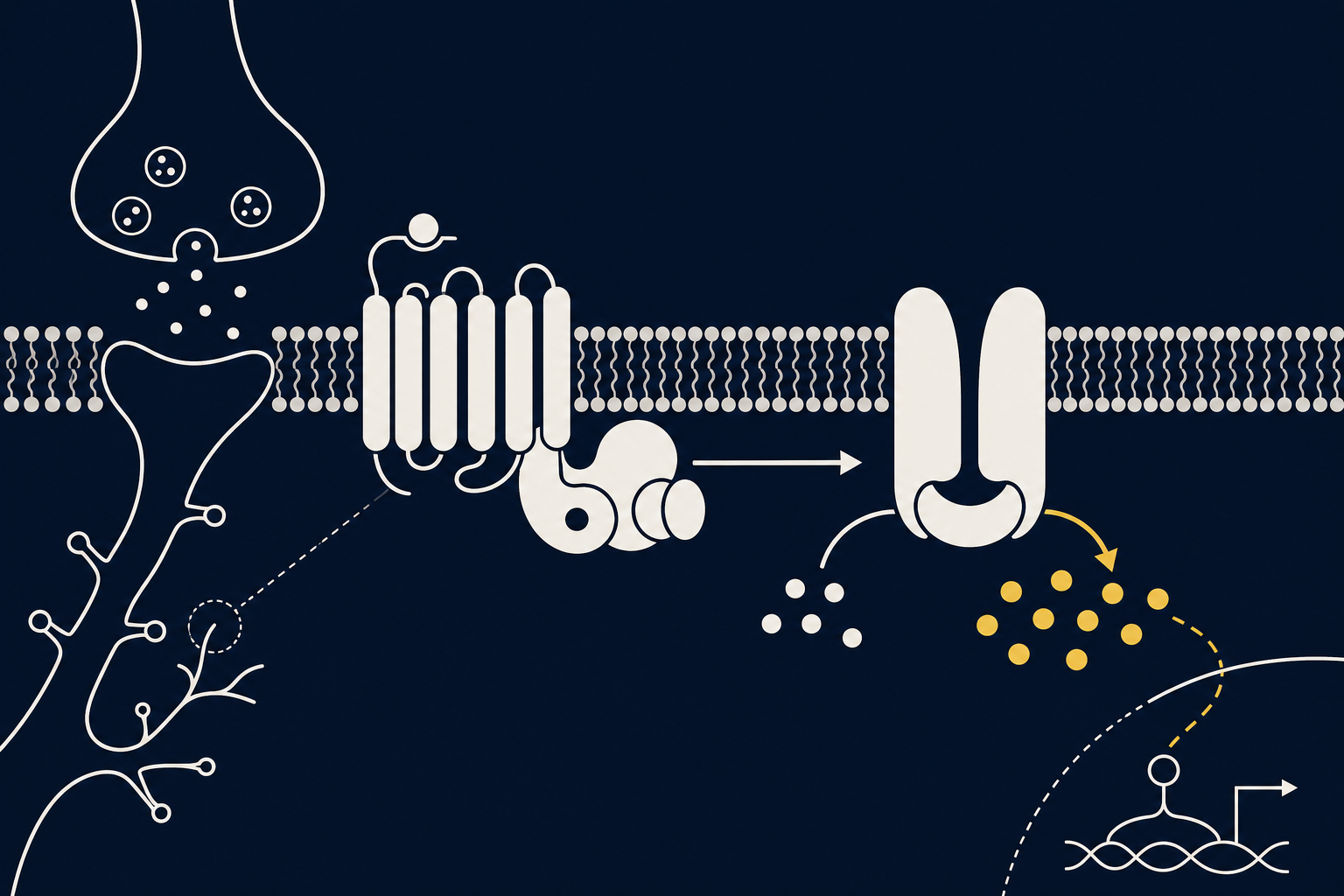
D1-рецептор (D1 Receptor) — метаботропный, сопряжённый с G-белком рецептор дофамина. У человека описано пять типов дофаминовых рецепторов — дофаминовые рецепторы D1–D5, которые делят на два семейства по типу G-белка и эффекту на клетку. D1-подобное семейство (D1 и D5) активирует аденилатциклазу и повышает цАМФ; D2-подобное семейство (D2, D3, D4), к которому относится D2-рецептор, наоборот, эту активность подавляет. D1-рецептор — самый многочисленный дофаминовый рецептор в ЦНС: он плотно представлен в стриатуме, префронтальной коре, гиппокампе и миндалине — структурах, формирующих мотивированное поведение, оценку ценности действия и удержание информации «в уме».
Как это работает
Молекулярный каскад D1-рецептора устроен как усилитель сигнала. Дофамин связывается с рецептором, тот меняет конформацию и активирует G-белок Gs, который стимулирует аденилатциклазу — фермент, превращающий АТФ в циклический аденозинмонофосфат (цАМФ). Рост концентрации цАМФ активирует протеинкиназу А (PKA), которая фосфорилирует множество мишеней, включая белок DARPP-32 — ключевой интегратор дофаминовых и глутаматных сигналов в шипиковых нейронах стриатума. Фосфорилированный DARPP-32 блокирует протеинфосфатазу-1, усиливая и продлевая эффект PKA — своего рода молекулярный переключатель, закрепляющий состояние нейрона.
В базальных ганглиях D1-рецепторы экспрессируются преимущественно на нейронах прямого пути — той группы клеток стриатума, которая при активации облегчает выбранное действие, в отличие от непрямого пути, где преобладает D2-рецептор. В префронтальной коре тот же каскад настраивает избирательность пирамидных нейронов, удерживающих активный след информации, — то есть саму рабочую память.
Особенность D1-сигнала в коре — не линейная, а колоколообразная, «инвертированная U-образная» зависимость: и слишком слабая, и слишком сильная стимуляция рецептора ухудшают работу нейронов, тогда как умеренный уровень активации обостряет настройку клеток и улучшает удержание информации.
Почему это важно для мотивации
D1-рецептор — точка, где биохимия дофамина превращается в мотивированное поведение. В стриатуме через прямой путь он усиливает сигнал «действие ведёт к ценному результату», подкрепляя выбор и инициацию усилия. В префронтальной коре тот же рецептор обеспечивает удержание цели в рабочей памяти достаточно долго, чтобы довести многошаговое действие до конца, — без этого намерение рассыпается на отвлечения. Поэтому колебания дофаминового тонуса заметно сказываются на способности начинать и доводить дела: недостаточная D1-стимуляция в коре связана с рассеянностью и слабым контролем импульсов, а избыточная или хронически завышенная — с ригидностью мышления и явлениями, которые описывает термин D1-перегрузка. Хроническая гиперстимуляция рецептора со временем запускает десенситизацию и даун-регуляцию рецепторов, из-за которых прежний уровень стимула перестаёт давать прежний эффект, и системе требуется всё более сильный стимул для того же чувства вовлечённости.
Что с этим делать
Управлять отдельным рецептором напрямую нельзя, но можно управлять условиями, в которых он работает. Поскольку эффективность D1-сигнала в коре описывается колоколообразной кривой, практический вывод — избегать крайностей: хронического недосыпа, снижающего дофаминовый тонус, и постоянной гиперстимуляции короткими вознаграждениями (лента уведомлений, азартные паттерны), которая толкает систему к перегрузке и последующей десенситизации. Достаточный сон, физическая нагрузка и разумные интервалы между сильными стимулами удерживают дофаминовую систему ближе к рабочему диапазону, в котором память и целенаправленное усилие функционируют лучше всего. Задачи, требующие устойчивого внимания, стоит разбивать на этапы с ощутимым промежуточным результатом — это опирается на естественную роль D1-рецептора в удержании цели.
Итог
D1-рецептор — главный «усилитель» дофаминового сигнала: через G-белок Gs и цАМФ он переводит короткий химический импульс в устойчивую перестройку возбудимости нейронов стриатума и префронтальной коры. Эта биохимия объясняет, почему мотивация и когнитивный контроль настолько чувствительны к балансу дофамина — и почему как дефицит, так и избыток стимуляции одинаково мешают эффективному, целенаправленному поведению.
Материал носит образовательный характер и не заменяет консультацию специалиста.