Кратко
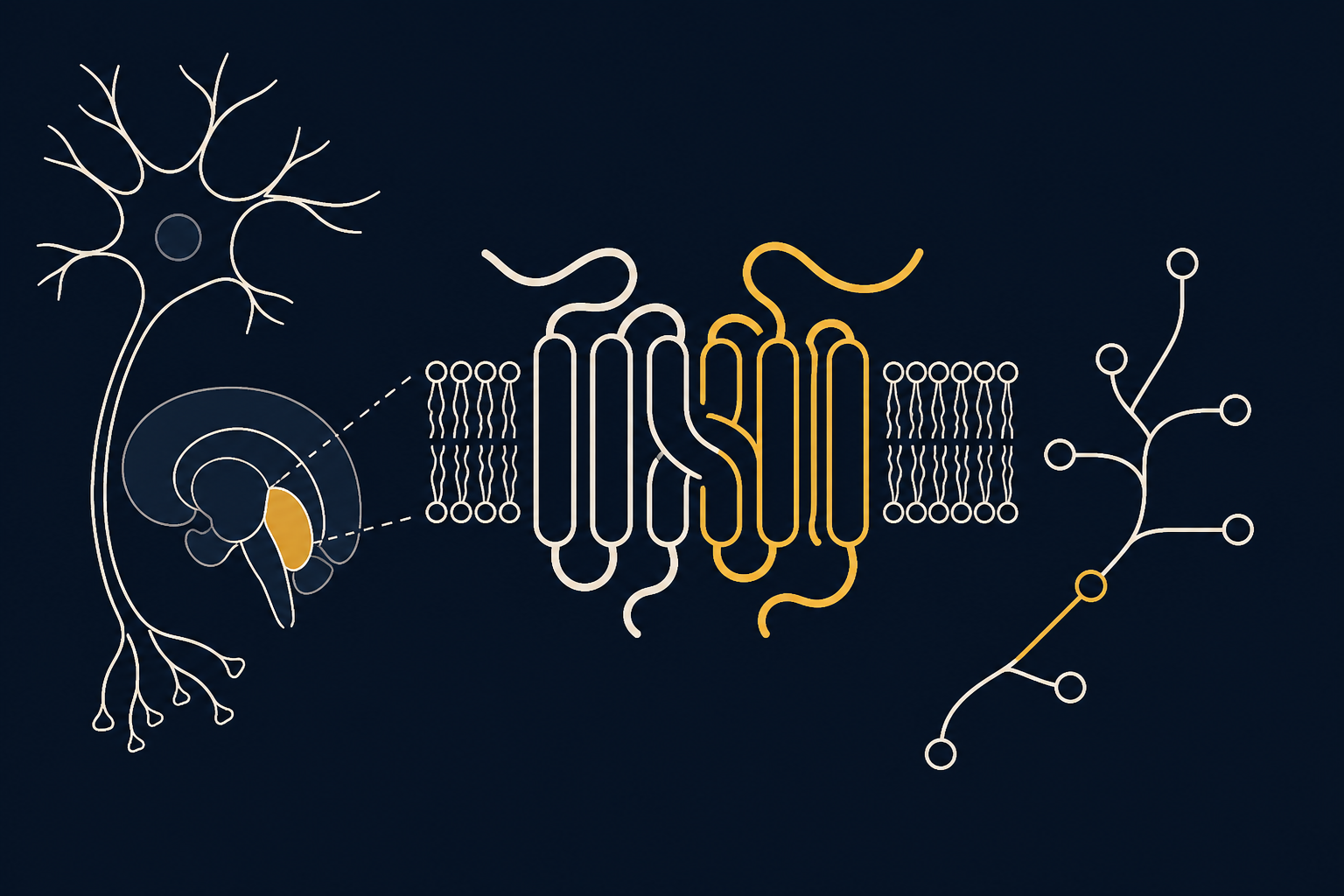
Рецепторы аденозина A2A — это молекулярный «тормоз» дофаминовой системы мозга. Они расположены на нейронах непрямого пути базальных ганглиев и физически сцеплены с дофаминовыми D2-рецепторами в единый функциональный комплекс — гетеромер. Когда аденозин активирует A2A, отклик D2-рецептора на дофамин слабеет; когда A2A блокируется — например, кофеином — дофаминовый сигнал, наоборот, усиливается. Эта пара рецепторов во многом определяет, готов ли организм в данный момент вкладывать усилие в достижение цели.
Что это такое
Аденозин — не классический нейромедиатор, а побочный продукт клеточного энергообмена: он накапливается по мере расходования АТФ и сигнализирует тканям об усталости. Его действие реализуется через четыре типа рецепторов, сопряжённых с G-белком (A1, A2A, A2B, A3), и именно A2A особенно плотно сконцентрирован в стриатуме — структуре, отвечающей за выбор и запуск действий. Несмотря на созвучное название, A2A-рецепторы не имеют отношения к α2A-адренорецепторам, которые реагируют на норадреналин и относятся к совершенно другому семейству. Общее представление о рецепторе как таковом даёт статья «A2A-рецептор»; здесь же рассматривается его специфическая роль в паре с дофаминовой системой.
Как это работает
На мембране стриатопаллидарных нейронов A2A- и D2-рецепторы образуют устойчивый гетеротетрамерный комплекс: два рецептора A2A соединены с двумя рецепторами D2 в единую сигнальную единицу. Внутри этого комплекса работает аллостерическое взаимодействие — активация A2A снижает сродство D2 к дофамину и ослабляет его внутриклеточный ответ, а блокада A2A восстанавливает полноценную работу D2. Сергей Ферре с коллегами показали, что именно эта архитектура, а не простое конкурентное вытеснение, объясняет, почему кофеин — неселективный антагонист аденозиновых рецепторов — усиливает дофаминовую передачу и субъективно ощущается как прилив бодрости и мотивации. Схожий механизм отчасти затрагивает и другие рецепторные пары мозга, изученные в статьях о 5-HT1A-рецепторе и его ауторецепторах, где регуляция также строится на тонком балансе двух систем.
Почему это важно для мотивации
Дофамин в прилежащем ядре и дорсальном стриатуме управляет не столько удовольствием, сколько готовностью прилагать усилие ради вознаграждения. Работы Джона Саламоне и коллег на животных моделях показали: блокада D2-рецепторов заставляет животных выбирать лёгкие, малозатратные варианты поведения вместо более трудных, но выгодных задач. Введение антагонистов A2A — избирательно и предсказуемо — обращает этот эффект вспять, возвращая готовность работать за более крупное вознаграждение. Иными словами, баланс активности A2A и D2 является одним из молекулярных переключателей между «мне лень» и «я готов постараться». Нарушение этого баланса рассматривается как один из механизмов патологической апатии, психомоторной заторможенности и части симптомов, сопровождающих депрессию и паркинсонизм.
Что с этим делать
Практический вывод из этой нейрохимии скромный, но конкретный: умеренное количество кофеина действительно способно временно повысить субъективную готовность к усилию за счёт блокады A2A-рецепторов, но это компенсаторный, а не ресурсный эффект — он не создаёт новую энергию, а лишь снимает тормоз с уже имеющейся дофаминовой активности. Злоупотребление кофеином истощает этот резерв быстрее и оборачивается снижением чувствительности рецепторов. Куда устойчивее работают факторы, которые поддерживают базовую дофаминовую регуляцию: достаточный сон (во сне аденозин выводится из тканей), регулярная физическая нагрузка и посильные, но значимые задачи, поддерживающие естественный цикл усилия и вознаграждения. Стоит также иметь в виду, что препараты, блокирующие D2-рецепторы вне головного мозга — в гипофизе, — могут вызывать гиперпролактинемию, ещё один пример того, как один и тот же рецептор влияет на разные системы организма.
Итог
Рецепторы аденозина A2A — это не самостоятельный «центр мотивации», а тонкий регулятор дофаминовой системы, встроенный с ней в единый молекулярный комплекс. Их работа определяет порог, начиная с которого мозг признаёт усилие оправданным, а блокада этих рецепторов кофеином — самый доступный и массовый способ временно сдвинуть этот порог. Понимание данного механизма помогает разумнее относиться и к кофеину, и к собственной готовности действовать в разные дни.
Материал носит образовательный характер и не заменяет консультацию специалиста.