Кратко
Протеинкиназа А (PKA, от англ. Protein Kinase A) — ферментный «исполнитель» дофаминового сигнала: она включается, когда в нейроне повышается концентрация циклического АМФ, и передаёт команду дальше, навешивая фосфатные группы на десятки других белков. Это один из главных механизмов, через которые кратковременный всплеск дофамина превращается в изменение возбудимости нейрона, экспрессии генов и, в конечном счёте, поведения. Без PKA сигнал от циклического АМФ остался бы просто скачком концентрации молекулы, ни на что не влияющим.
Что это
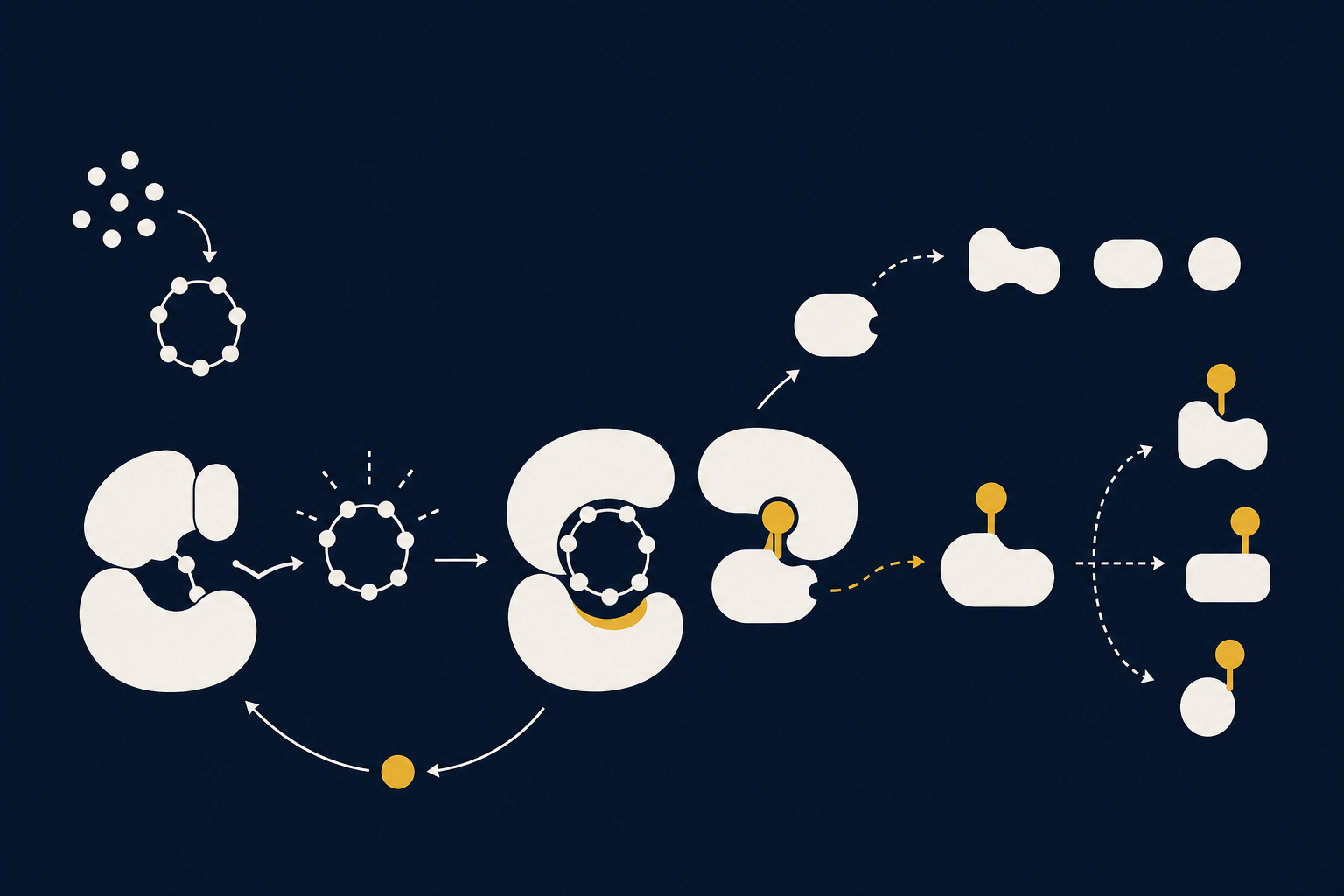
PKA — это цАМФ-зависимая протеинкиназа, фермент из семейства серин/треониновых киназ. В неактивном состоянии молекула существует как холоэнзим — тетрамер из двух регуляторных (R) и двух каталитических (C) субъединиц, где регуляторные субъединицы блокируют каталитические. Как только к каждой из регуляторных субъединиц присоединяется по две молекулы цАМФ, комплекс меняет конформацию и высвобождает каталитические субъединицы — именно они переносят фосфат с АТФ на белки-мишени. Это подробно описано в структурных обзорах Сюзан Тейлор и коллег, посвящённых архитектуре PKA как модели аллостерической регуляции киназ.
В нейрохимии мотивации PKA занимает центральное место в D1-каскаде: дофамин → D1-рецептор → аденилатциклаза → циклический АМФ → PKA → фосфорилирование мишеней. Это один из узлов каскада, где сигнал одновременно расходится на быстрые электрофизиологические эффекты (изменение ионных токов) и медленные — изменение транскрипции генов.
Как это работает
Активированная PKA фосфорилирует три больших класса субстратов. Во-первых, ионные каналы и рецепторы на мембране — это меняет возбудимость нейрона в течение секунд. Во-вторых, регуляторный белок DARPP-32: фосфорилирование по треонину-34 превращает его в мощный ингибитор фосфатазы PP-1, что резко усиливает и продлевает эффект самой PKA — молекулярный усилитель обратной связи, описанный в работах Пола Грингарда и коллег. В-третьих, транскрипционный фактор CREB: фосфорилированный по серину-133, он запускает CRE-зависимые гены, лежащие в основе долговременных изменений синапсов и памяти (обзор Эрика Канделя, 2012).
Важная деталь: PKA не «плывёт» свободно по клетке — якорные белки AKAP удерживают её холоэнзим рядом с конкретными субстратами, поэтому один и тот же фермент даёт разные эффекты в разных компартментах нейрона в зависимости от того, где он закреплён.
Почему это важно для мотивации
PKA — это точка, где химический сигнал дофамина конвертируется в изменение поведения. Активация D1-рецепторов на стриатальных нейронах через PKA и DARPP-32 усиливает отклик на возбуждающий вход и облегчает формирование связи «усилие → вознаграждение». Именно этот каскад объясняет, почему предвкушение награды повышает готовность действовать: без исправно работающей PKA дофаминовый всплеск не транслируется в устойчивое изменение возбудимости нейронов прилежащего ядра.
Обратная сторона в том, что каскад цАМФ→PKA легко перегрузить: хронически повышенный тонус дофаминовой и норадренергической систем — состояние, которое описывается как хронический возбудительный стресс — держит PKA в постоянно повышенной активности. Результат — не усиление мотивации, а десенситизация: рецепторы и ферменты каскада адаптируются к постоянной стимуляции, и для того же эффекта требуется всё больший сигнал. Это одна из молекулярных основ феномена, когда привычные стимулы — работа, соцсети, еда — перестают вызывать прежний отклик.
Что с этим делать
Напрямую «включить» или «выключить» PKA доступными средствами нельзя и не нужно — это фундаментальный фермент, вмешательство в который чревато системными последствиями. Но логику каскада можно учитывать на уровне образа жизни. Каскаду цАМФ→PKA нужны кофакторы: например, магний участвует в регуляции аденилатциклазы и АТФ-зависимых ферментов, и его хронический дефицит теоретически может смещать баланс каскада. Более практичный вывод — избегать хронической гиперстимуляции дофаминовой системы (постоянные уведомления, стимуляторы, хаотичный сон), которая держит цАМФ-каскад в перевозбуждении и способствует десенситизации. Регулярное чередование нагрузки и восстановления даёт каскаду время вернуться к базовому уровню между всплесками активации — судя по имеющимся данным, именно колебание, а не постоянно высокий тонус, поддерживает чувствительность системы вознаграждения.
Итог
Протеинкиназа А — молекулярный переводчик, который превращает всплеск цАМФ в конкретное действие: изменение ионных токов, активацию DARPP-32, включение генов через CREB. Это ключевое звено D1-каскада дофаминовой системы, без которого сигнал о награде не превращался бы в устойчивое изменение поведения. Понимание этого механизма объясняет не только то, как работает мотивация, но и то, почему хроническая гиперстимуляция притупляет чувствительность к вознаграждению.
Материал носит образовательный характер и не заменяет консультацию специалиста.