Кратко
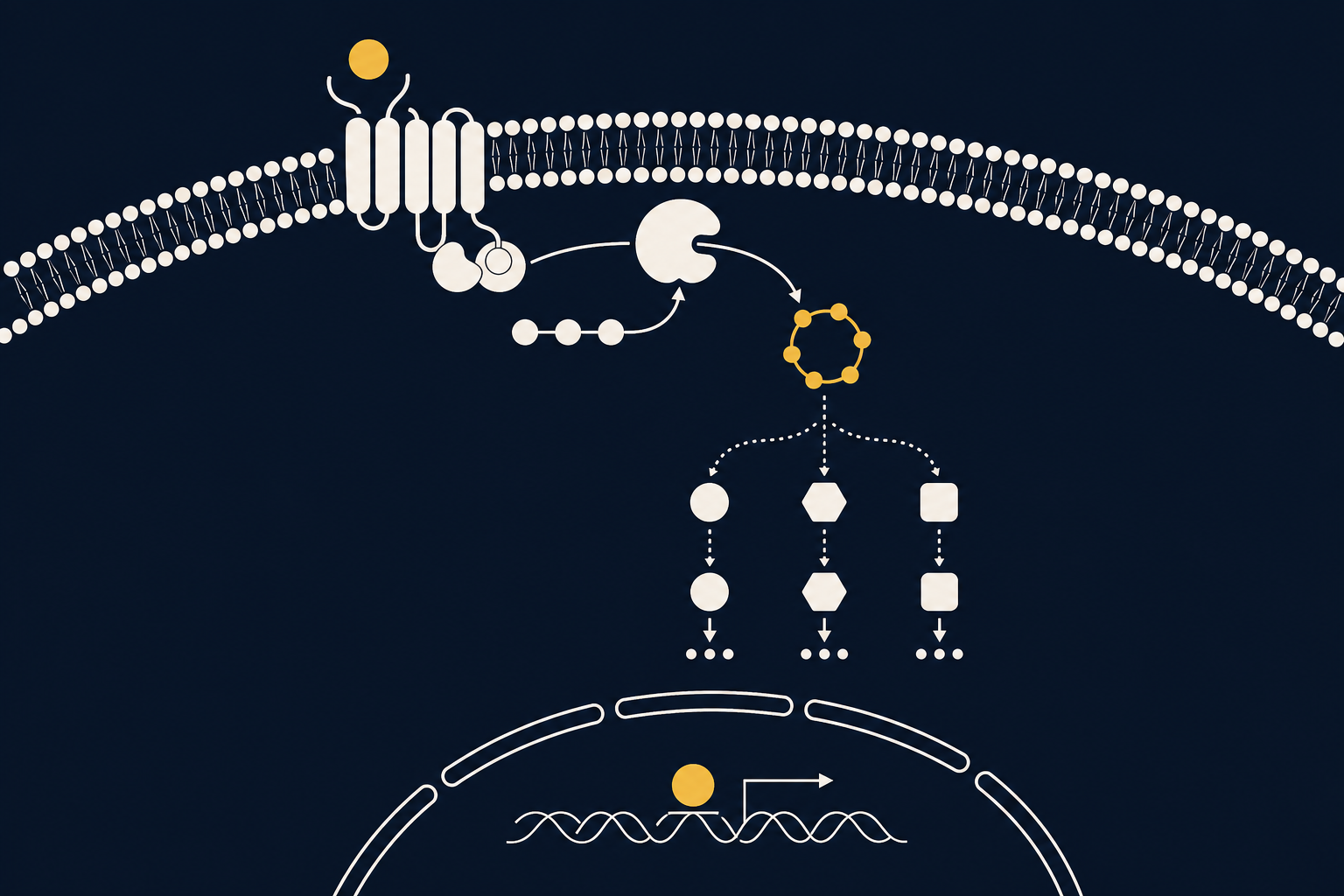
Циклический аденозинмонофосфат, или цАМФ (cAMP), — универсальный внутриклеточный посредник, через который дофаминовые рецепторы передают сигнал внутрь нейрона. Когда дофамин активирует D1-рецептор, концентрация цАМФ растёт и запускает возбуждающий каскад; когда сигнал идёт через D2-рецептор, синтез цАМФ, напротив, подавляется. Этот простой переключатель «больше цАМФ / меньше цАМФ» лежит в основе того, как дофамин настраивает готовность нейрона к работе.
Что это
цАМФ — небольшая циклическая молекула, образующаяся из АТФ. Её открыл в 1950-х годах американский биохимик Эрл Сазерленд, изучая, как гормоны вроде адреналина заставляют печень расщеплять гликоген; за описание цАМФ как «второго посредника» он получил Нобелевскую премию по физиологии и медицине 1971 года. Идея оказалась универсальной: гормон или нейромедиатор (первый посредник) не проникает в клетку, а связывается с рецептором на её поверхности, и уже внутри клетки его сигнал транслируется через изменение концентрации цАМФ (второй посредник). В нейрохимии мотивации цАМФ — молекулярное звено, превращающее внеклеточный сигнал дофамина в конкретные внутриклеточные события: фосфорилирование белков, изменение возбудимости мембраны и, в конечном счёте, экспрессию генов.
Как это работает
Дофаминовые рецепторы относятся к G-белоксвязанным рецепторам и напрямую не открывают ионные каналы. D1- и D5-рецепторы сопряжены с G-белком Gs/olf, который активирует фермент аденилатциклазу: она превращает АТФ в цАМФ, и его концентрация в клетке возрастает. D2-, D3- и D4-рецепторы сопряжены с G-белком Gi/o, который, напротив, тормозит аденилатциклазу и снижает уровень цАМФ. Дальнейшая судьба сигнала зависит от одной ключевой мишени — протеинкиназы А (PKA): цАМФ связывается с её регуляторными субъединицами и высвобождает каталитические, которые фосфорилируют десятки белков-мишеней. Одна из важнейших — DARPP-32, которая в фосфорилированном виде блокирует фосфатазу PP-1 и тем самым продлевает и усиливает эффект PKA. При достаточно сильной и длительной активации PKA проникает и в ядро клетки, где фосфорилирует транскрипционный фактор CREB — это уже не секунды, а часы и дни: изменённая экспрессия генов лежит в основе долговременной синаптической пластичности и памяти.
Почему это важно для мотивации
Каскад цАМФ — молекулярный переводчик между событием «дофамин выделился» и результатом «нейрон изменил своё поведение». Умеренный, своевременный подъём цАМФ через D1-путь в префронтальной коре помогает удерживать цель в рабочей памяти и подавлять отвлекающие сигналы — именно так выглядит собранность и способность прилагать усилие. Но избыточная, хроническая D1-стимуляция — состояние, которое описывают как D1-перегрузку, — доводит PKA-сигнал до неселективного, и он начинает мешать той же рабочей памяти. Постоянно повышенный фон цАМФ и PKA-активности характерен и для более общей картины — хронического возбудительного стресса, при котором система «застревает» на высокой передаче сигнала и хуже возвращается к спокойному базовому уровню. Для мотивации важен не абсолютный уровень цАМФ, а его способность вовремя подниматься и вовремя снижаться.
Что с этим делать
Напрямую управлять концентрацией цАМФ невозможно и не нужно — это внутриклеточный процесс, а не объект сознательного контроля. Но есть косвенные факторы, задающие фон, на котором каскад работает предсказуемо. Достаточный сон и устойчивый режим дня снижают вероятность хронической гиперактивации дофаминовых путей. Разумно не рассчитывать на пик когнитивной собранности одновременно с пиком острого стресса или тревоги — в этот момент D1–цАМФ–PKA-сигнал уже близок к избыточному, и дополнительное давление или жёсткий дедлайн лишь толкают систему дальше. Отдельный фактор общей нейрохимической устойчивости — достаточный статус магния, который естественным образом ограничивает часть механизмов повышения возбудимости мембраны. Это не способы «повысить цАМФ», а способы не доводить систему до перегрузки, при которой сигнал теряет избирательность.
Итог
цАМФ — один из самых фундаментальных вторичных посредников в биологии клетки и ключевое звено дофаминовой сигнализации: его рост при D1-активации и снижение при D2-активации определяют, как внеклеточный сигнал дофамина превращается во внутриклеточные события — от кратковременного фосфорилирования белков через PKA и DARPP-32 до долговременных изменений экспрессии генов через CREB. Понимание этого каскада объясняет, почему дофаминовый сигнал не работает по принципу «больше — всегда лучше»: то, что делает нейрон собранным при умеренной активации, при избыточной превращается в помеху.
Материал носит образовательный характер и не заменяет консультацию специалиста.