Кратко
β-аррестин 2 — внутриклеточный белок-адаптер, который садится на активированные G-белок-сопряжённые рецепторы (GPCR), в том числе дофаминовые, и «выключает» их классический сигнал. Но у этого белка есть вторая, менее очевидная роль: он сам становится платформой для нового каскада — Akt/GSK3, — который влияет на дофамин-зависимое поведение. Иными словами, один и тот же белок сначала гасит один сигнал, а затем запускает другой.
Что это
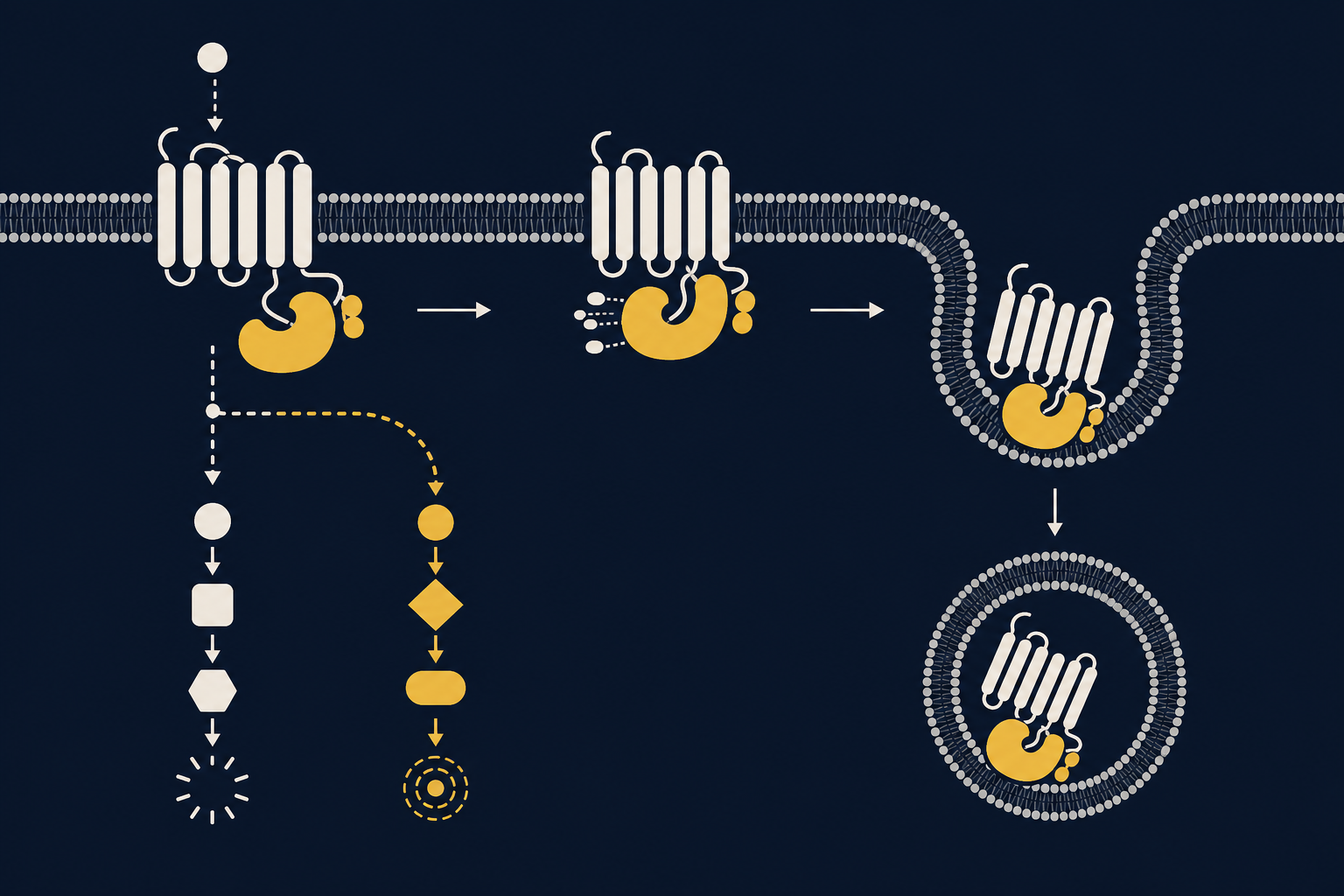
β-аррестин 2 (β-Arrestin 2, ARRB2) относится к семейству аррестинов — небольших цитоплазматических белков, открытых как регуляторы фоторецептора родопсина, а затем найденных практически у всех GPCR, включая рецепторы дофамина, серотонина и аденозина. Классическая функция аррестинов — десенситизация: после того как рецептор активирован и фосфорилирован киназами GRK, аррестин связывается с ним, блокирует дальнейшее взаимодействие с G-белком и запускает интернализацию рецептора внутрь клетки через эндоцитоз.
Долгое время аррестины считали просто «стоп-краном» сигнала. Ситуация изменилась, когда выяснилось, что аррестины умеют собирать вокруг себя собственные сигнальные комплексы — киназы, фосфатазы, компоненты MAPK-каскада — и превращаться в самостоятельный передатчик информации, независимый от G-белка. Этот сдвиг во взглядах подробно описан в обзоре Лефковица и Шеноя, поставившем аррестины в один ряд с G-белками как полноценных трансдьюсеров рецепторного сигнала.
Как это работает
У дофаминовых рецепторов, особенно D1-рецептора и рецепторов D2-класса, есть два параллельных режима работы. Первый — классический, через G-белок и аденилатциклазу, меняющий уровень цАМФ. Второй — «аррестиновый»: дофамин способствует формированию тройного комплекса из киназы Akt, β-аррестина 2 и фосфатазы PP2A. В этом комплексе PP2A дефосфорилирует и тем самым инактивирует Akt, что снимает тормоз с киназы GSK3 и повышает её активность. Ключевое наблюдение здесь в том, что этот путь не зависит от цАМФ: у мышей с удалённым геном β-аррестина 2 классическая цАМФ-сигнализация сохраняется, а вот регуляция Akt/GSK3 дофамином полностью исчезает.
Такое избирательное включение одной ветки сигнала при сохранении другой называют смещённой (biased) сигнализацией: один и тот же рецептор, в зависимости от лиганда и контекста, может «тянуть одеяло» либо к G-белку, либо к аррестину. Похожий принцип регуляции через фосфорилирование, аррестин-связывание и интернализацию описан и для серотониновых рецепторов — например, 5-HT1A-рецептора и 5-HT2A-рецептора, а также для аденозинового A2A-рецептора, который образует комплексы с дофаминовыми рецепторами в стриатуме.
Почему это важно для мотивации
Путь Akt/GSK3 напрямую связан с дофамин-зависимым поведением: у мышей, лишённых β-аррестина 2, снижена локомоторная и поведенческая реакция на дофаминергические стимулы, а фармакологические эффекты, зависящие от повышенного дофамина, ослабевают. Поскольку дофамин — центральный нейромедиатор системы вознаграждения и целенаправленного усилия, любое изменение баланса между G-белковой и аррестиновой ветвями сигнала способно менять то, насколько охотно человек инициирует действие, насколько устойчива его мотивация и как он реагирует на вознаграждение или его отсутствие.
Этот же механизм оказался важен для психофармакологии: снижение активности Akt через данный путь связывают с действием антипсихотиков и стабилизаторов настроения, а нарушения баланса Akt/GSK3 обсуждаются как один из молекулярных факторов, способствующих ангедонии и снижению побудительной активности при некоторых психических расстройствах.
Что с этим делать
Напрямую «настроить» собственный β-аррестин 2 нельзя — это не тот уровень, на котором работают повседневные привычки. Но полезно иметь в виду сам принцип: дофаминовые (и в целом GPCR-опосредованные) эффекты не сводятся к простому «больше дофамина — больше мотивации». Существует вторая, независимая от цАМФ линия регуляции, которая формирует более медленные и устойчивые изменения в клетке. Именно на неё нацелены некоторые современные препараты, разрабатываемые как «смещённые» (biased) лиганды — молекулы, избирательно активирующие один сигнальный путь рецептора и по возможности не задевающие другой, что в перспективе может снижать побочные эффекты классической дофаминергической терапии.
Итог
β-аррестин 2 — пример того, как один и тот же белок способен быть одновременно «выключателем» и «переключателем» сигнала рецептора. Через комплекс с Akt и PP2A он формирует альтернативный, независимый от цАМФ канал регуляции дофаминовых эффектов, значимый для поведения, мотивации и фармакологии психических расстройств. Это подчёркивает общий принцип современной нейрохимии: сигнал рецептора — это не одна линия, а сеть параллельных путей, и именно их баланс определяет итоговый эффект нейромедиатора.
Материал носит образовательный характер и не заменяет консультацию специалиста.